

- 最新进展
- 产品信息
- 参考文献
MHC四聚体
在免疫学研究领域,精准识别和量化抗原特异性T细胞是解析免疫应答机制、开发新型疫苗和免疫治疗策略的核心环节。MHC四聚体技术自1996年诞生以来,凭借其高灵敏度、高特异性和单细胞水平分析能力,已成为免疫监测与治疗领域的“金标准”。
这项技术通过将MHC分子与抗原肽结合形成四聚体结构,结合荧光标记和流式细胞术,实现了对抗原特异性T细胞的精准捕捉与量化分析,为免疫学研究带来了革命性突破。
技术原理:四聚体结构的创新设计
MHC四聚体的核心在于其独特的分子结构。MHC分子在免疫系统中负责将抗原肽呈递给T细胞,而MHC四聚体技术通过生物素-链霉亲和素系统,将四个MHC-肽复合物连接成四聚体结构。这种设计显著提高了MHC-肽复合物与T细胞受体(TCR)的结合亲和力和稳定性,解决了传统MHC-肽单体亲和力低、解离速度快的问题。四聚体结构中的每个MHC分子均可与T细胞表面的TCR结合,形成多价相互作用,从而实现对低频抗原特异性T细胞的精准检测。
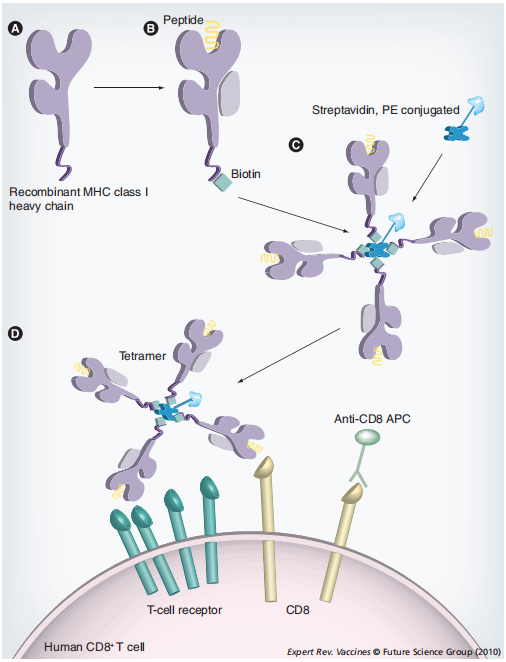
MHC四聚体的构建通常涉及以下步骤:抗原肽制备-生物素化-四聚化-荧光标记
MHC四聚体的优势
相较于酶联免疫斑点检测(ELISPOT)、胞内细胞因子染色(ICS)等传统用于 T 细胞检测的方法,MHC 四聚体优势如下:
高特异性:直接且精准地识别抗原肽特异性的 CD8⁺T 细胞表面的TCR
高敏感性:大量实验数据有力地证实,其具有很高的亲和力,和很低的本底背景信号
操作简单便捷:无需对 T 细胞进行抗原肽刺激以及扩增培养等操作
重复性高:检测结果的重复性十分出色
从基础研究到临床转化
MHC四聚体技术在多个领域展现出广泛应用价值,成为免疫学研究的“瑞士军刀”。
肿瘤免疫治疗
在肿瘤免疫治疗中,MHC四聚体技术被用于识别和分离肿瘤特异性T细胞,为个性化治疗提供关键数据。主要应用于新生抗原/表位肽疫苗/DC疫苗/CTL细胞回输/TCR-T疗法等。
例如,在黑色素瘤新抗原疫苗的研究中,研究人员通过MHC四聚体技术筛选出能够激活T细胞免疫应答的抗原肽,并结合单细胞测序技术分析接种疫苗前后的T细胞基因表达图谱。结果显示,个性化新抗原疫苗能够诱导强烈的T细胞反应,证明了其在临床应用中的安全性和有效性。
传染病研究
MHC四聚体技术在传染病(新冠病毒/流感病毒/巨细胞病毒/艾滋病毒/EB病毒/HPV等)研究中同样发挥重要作用。例如,在SARS-CoV-2的研究中,研究人员利用MHC四聚体技术分析了感染者鼻粘膜组织中的抗原特异性CD8+ T细胞,揭示了鼻腔驻留T细胞的免疫应答机制。此外,该技术还被用于HIV、流感病毒等病原体的抗原表位筛选,为疫苗开发提供了重要依据。
自身免疫疾病
在自身免疫疾病(糖尿病/过敏/类风湿性关节炎)中,MHC四聚体技术可用于精确定位负责病理免疫反应的特异性T细胞。例如,在1型糖尿病的研究中,研究人员通过MHC四聚体技术识别出针对胰岛β细胞的自身反应性T细胞,为开发靶向治疗策略提供了可能。
移植免疫
在器官移植领域,MHC四聚体技术可用于监测移植受者的免疫应答,评估排斥反应风险。通过检测供体特异性T细胞的频率和表型,医生可以及时调整免疫抑制方案,提高移植成功率。
市场前景:免疫监测与治疗的核心工具
随着免疫学研究的深入和个性化医疗的兴起,MHC四聚体技术的市场需求持续增长。据市场调研机构预测,全球MHC四聚体市场规模将在2031年达到20.49百万美元,年复合增长率为7.9%。北美地区凭借其强大的医药研发实力占据最大市场份额,而亚太地区则因人口基数大、科研投入增加,成为增长最快的区域。
展望未来,MHC四聚体技术的发展将聚焦于多重检测、自动化和计算分析的创新。多重检测技术允许在单个样本中同时检测多种T细胞特异性,从而全面分析免疫反应。自动化技术则通过标准化流程和提高通量,减少实验室间的差异,提高实验可重复性。此外,计算分析工具的进步将帮助研究人员处理MHC四聚体实验生成的大型数据集,深入解析免疫系统动力学和疫苗疗效。
MHC-I Tetramer部分热销产品清单(更多请咨询我们):
|
病原/肿瘤 |
产品名称 |
抗原 |
序列 |
MHC |
位置 |
货号 |
|
EBV |
HLA-A*0201/YLLEILWRL-PE Labelled Tetramer |
EBV.LMP1 |
YLLEILWRL |
HLA-A*0201 |
125~133 |
UA089001 |
|
HLA-A*0201/YLQQNWWTL-PE Labelled Tetramer |
EBV.LMP1 |
YLQQNWWTL |
HLA-A*0201 |
159~167 |
UA089003 |
|
|
HPV |
H-2D(b)/RAHYNIVTF-PE Labelled Tetramer |
HPV16E7 |
RAHYNIVTF |
H-2D(b) |
49~57 |
UA089002 |
|
H-2K(b)/EVYDFAFRDL-PE Labelled Tetramer |
HPV16.E6 |
EVYDFAFRDL |
H-2K(b) |
48~57 |
UA089004 |
|
|
HLA-A0201/KLPDLCTEL-PE Labelled Tetramer |
HPV18.E6 |
KLPDLCTEL |
HLA-A0201 |
13~21 |
UA089005 |
|
|
HLA-A0201/KLTNTGLYNL-PE Labelled Tetramer |
HPV18.E6 |
KLTNTGLYNL |
HLA-A0201 |
92~101 |
UA089006 |
|
|
HLA-A0201/TLQDIVLHL-PE Labelled Tetramer |
HPV18.E7 |
TLQDIVLHL |
HLA-A0201 |
7~15 |
UA089007 |
|
|
HLA-A0201/QLFLNTLSFV-PE Labelled Tetramer |
HPV18.E7 |
QLFLNTLSFV |
HLA-A0201 |
88~97 |
UA089008 |
|
|
HLA-A1101/GVNHQHLPAR-PE Labelled Tetramer |
HPV18.E7 |
GVNHQHLPAR |
HLA-A1101 |
43~52 |
UA089009 |
|
|
Influenza A virus |
H-2D(b)/ASNENMETM-PE Labelled Tetramer |
Flu.NP |
ASNENMETM |
H-2D(b) |
366~374 |
UA089010 |
|
H-2K(d)/TYQRTRALV-PE Labelled Tetramer |
Flu NP |
TYQRTRALV |
H-2K(d) |
147~155 |
UA089011 |
|
|
H-2D(b)/ASNENMDTM-PE Labelled Tetramer |
Flu.NP |
ASNENMDTM |
H-2D(b) |
366~374 |
UA089012 |
|
|
LCMV |
H-2D(b)/KAVYNFATM-PE Labelled Tetramer |
GP 33 |
KAVYNFATM |
H-2D(b) |
33~41 |
UA089013 |
|
H-2D(b)/FQPQNGQFI-PE Labelled Tetramer |
LCMV NP |
FQPQNGQFI |
H-2D(b) |
396~404 |
UA089014 |
|
|
肿瘤相关抗原 |
HLA-A*1101/VVVGADGVGK-PE Labelled Tetramer |
KRAS |
VVVGADGVGK |
HLA-A*1101 |
7~16 |
UA089015 |
|
HLA-A*1101/VVVGAVGVGK-PE Labelled Tetramer |
KRAS |
VVVGAVGVGK |
HLA-A*1101 |
7~16 |
UA089016 |
|
|
HLA-A*0201/KLVVVGAVGV-PE Labelled Tetramer |
KRAS |
KLVVVGAVGV |
HLA-A*0201 |
5~14 |
UA089017 |
|
|
HLA-A*0201/SLLMWITQC-PE Labelled Tetramer |
NY-ESO1 |
SLLMWITQC |
HLA-A*0201 |
157~165 |
UA089018 |
|
|
HLA-A*0201/LMWITQCFL-PE Labelled Tetramer |
NY-ESO2 |
LMWITQCFL |
HLA-A*0201 |
159~167 |
UA089019 |
|
|
H-2D(b)/RMFPNAPYL-PE Labelled Tetramer |
WT1 |
RMFPNAPYL |
H-2D(b) |
126~134 |
UA089020 |
|
|
HLA-A*0201/CMTWNQMNL-PE Labelled Tetramer |
WT2 |
CMTWNQMNL |
HLA-A*0201 |
235~243 |
UA089021 |
|
|
HLA-A*1101/KTCQRKFSR-PE Labelled Tetramer |
WT3 |
KTCQRKFSR |
HLA-A*1101 |
386~394 |
UA089022 |
|
|
模式抗原 |
H-2K(b)/SIINFEKL -PE Labelled Tetramer |
OVA |
SIINFEKL |
H-2K(b) |
257~264 |
UA089023 |

扫码查看更多~
参考文献:
- Donahoe SM, Moretto WJ, Samuel RV et al. Direct measurement of CD8+T cell responses in macaques infected with simian immunodeficiency virus. Virology (2000).
- Bakker AH, Hoppes R, Linnemann C et al.Conditional MHC class I ligands and peptide exchange technology for the human MHC gene products HLA-A1, -A3, -A11, and -B7. Proc. Natl Acad. Sci. USA(2008).
- Stuart Sims; Christian B. Willberg; Paul Klenerman. MHC–peptide tetramers for the analysis ofantigen-specific T cells. Expert Review of Vaccines(2010).
- Asbjørn Christophersen. Peptide-MHC class I and class II tetramers: From flowto mass cytometry. HLA(2019).
声明:本篇文章在创作中部分采用了人工智能辅助。如有任何内容涉及版权或知识产权问题,敬请告知,我们承诺将在第一时间核实并撤下。
买重组蛋白,找南京优爱
优爱蛋白专注于提供药物研发、细胞治疗、基因治疗、基础科研所需各种蛋白类试剂原材料和服务,包括药物靶点蛋白、免疫检查点蛋白、细胞因子、工具酶、 蛋白定制表达、全长跨膜蛋白开发等。优爱致力于为客户提供优质的产品和专业服务,打造具有国际竞争力的高新技术企业。

- Donahoe SM, Moretto WJ, Samuel RV et al. Direct measurement of CD8+T cell responses in macaques infected with simian immunodeficiency virus. Virology (2000).
- Bakker AH, Hoppes R, Linnemann C et al.Conditional MHC class I ligands and peptide exchange technology for the human MHC gene products HLA-A1, -A3, -A11, and -B7. Proc. Natl Acad. Sci. USA(2008).
- Stuart Sims; Christian B. Willberg; Paul Klenerman. MHC–peptide tetramers for the analysis ofantigen-specific T cells. Expert Review of Vaccines(2010).
- Asbjørn Christophersen. Peptide-MHC class I and class II tetramers: From flowto mass cytometry. HLA(2019).










