

- 最新进展
- 产品信息
MHC-Ib四聚体
MHC I类分子通过特异性呈递机制展示内源性抗原衍生肽段,其核心功能在于将细胞质内产生的8-11个氨基酸长度的短肽(如病毒感染相关病毒肽或肿瘤相关抗原肽)呈递至细胞毒性T淋巴细胞(CTL)表面。这一生物学过程包含三个精密调控的步骤:首先,蛋白酶体系统通过泛素-蛋白酶体途径对异常蛋白进行特异性切割,生成符合长度要求的抗原肽;其次,抗原加工相关转运体(TAP)通过异源二聚体结构将胞质溶胶中的抗原肽主动转运至内质网腔;最终,在内质网分子伴侣(如钙联蛋白)辅助下,抗原肽与MHC I类分子的α1/α2结构域形成稳定复合物,经高尔基体网络转运至细胞膜表面,构成TCR识别及免疫监视的分子基础。
尽管MHC I类与II类分子均属于免疫球蛋白超家族并共享抗原呈递功能,两者在抗原来源、细胞定位及免疫应答调控层面呈现显著差异。在深入解析MHC I类分子作用机制后,我们将聚焦MHC II类分子系统——这类分子专职处理外源性抗原(如胞外病原体蛋白),通过内体-溶酶体途径加工形成13-25个氨基酸的长肽,并特异性激活CD4+ T辅助细胞,从而在适应性免疫应答的启动与调控中发挥核心枢纽作用。
MHC II类分子
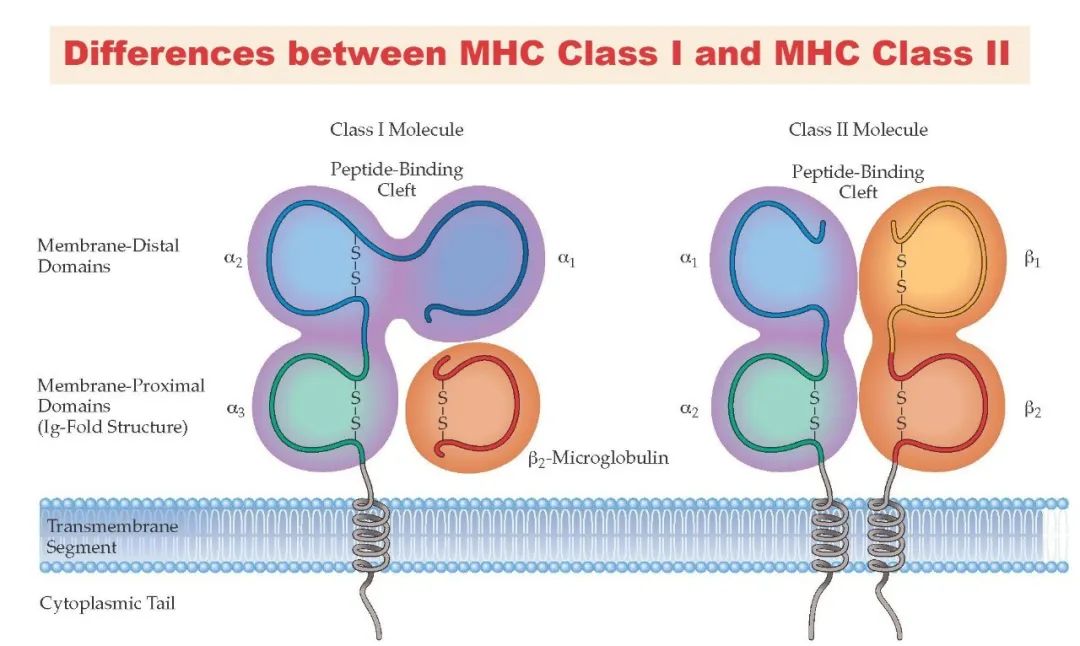
MHC II类分子由人类第6号染色体HLA-D区基因编码,其分子系统表现出与MHC I类分子同源的遗传多态性特征——不同个体间MHC II类分子的等位基因变异直接导致抗原呈递谱系的差异。值得注意的是,MHC II类分子的肽结合沟槽呈现独特的开放型构象:相较于MHC I类分子两端封闭的抗原结合裂隙,MHC II类分子的结合沟槽在两端呈开放状态,这种结构特性允许其容纳长度为13-25个氨基酸的延伸型抗原肽段(远超MHC I类分子8-11个氨基酸的短肽结合容量)。该结构-功能适应性使MHC II类分子能够有效呈递外源性蛋白酶体加工产生的较长肽段,为CD4+ T辅助细胞提供更丰富的表位识别界面。
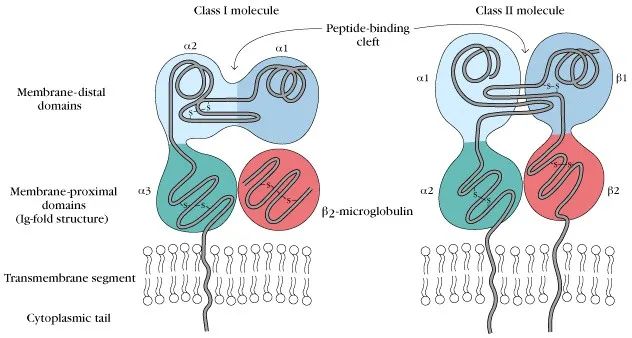
在深入解析MHC II类分子介导的抗原呈递机制前,我们首先对比MHC I类分子与MHC II类分子的核心功能差异:MHC I类分子通过交叉呈递机制识别内源性抗原(如病毒感染相关蛋白或肿瘤突变蛋白),并将8-11个氨基酸的短肽精准呈递至细胞毒性T淋巴细胞(CTL)的TCR识别界面;而MHC II类分子则专职处理外源性抗原(如胞外寄生菌或真菌蛋白),通过内体-溶酶体途径加工形成13-25个氨基酸的延伸肽段,最终激活CD4+ T辅助细胞(Th细胞)。
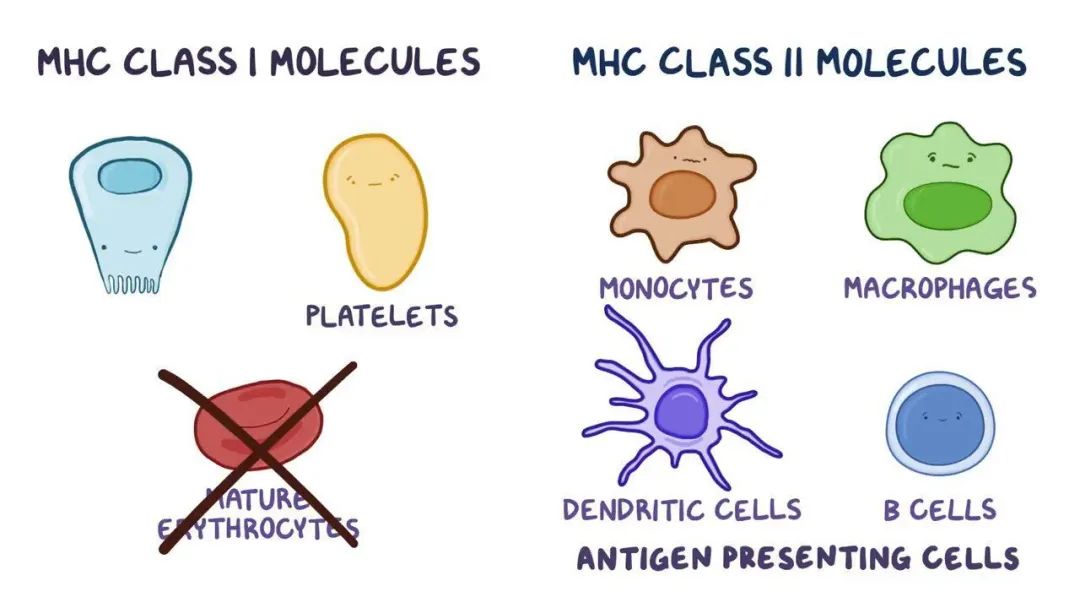
从细胞分布特征分析,MHC I类分子构成性表达于几乎所有有核细胞表面,形成覆盖全身的免疫监视网络——这种广泛表达模式使CTL能够实时检测全身各组织细胞的蛋白合成状态,有效遏制病毒感染或肿瘤转化;而MHC II类分子的表达严格局限于专职抗原呈递细胞(包括B淋巴细胞、单核吞噬细胞及树突状细胞),其功能定位如同"环境监测哨卡"——通过选择性抽样局部微环境中的外源性蛋白信息,将危险信号级联放大并传递至T细胞应答网络。这种差异化表达策略既保障了免疫监视的全面性,又避免了非特异性免疫激活导致的组织损伤。
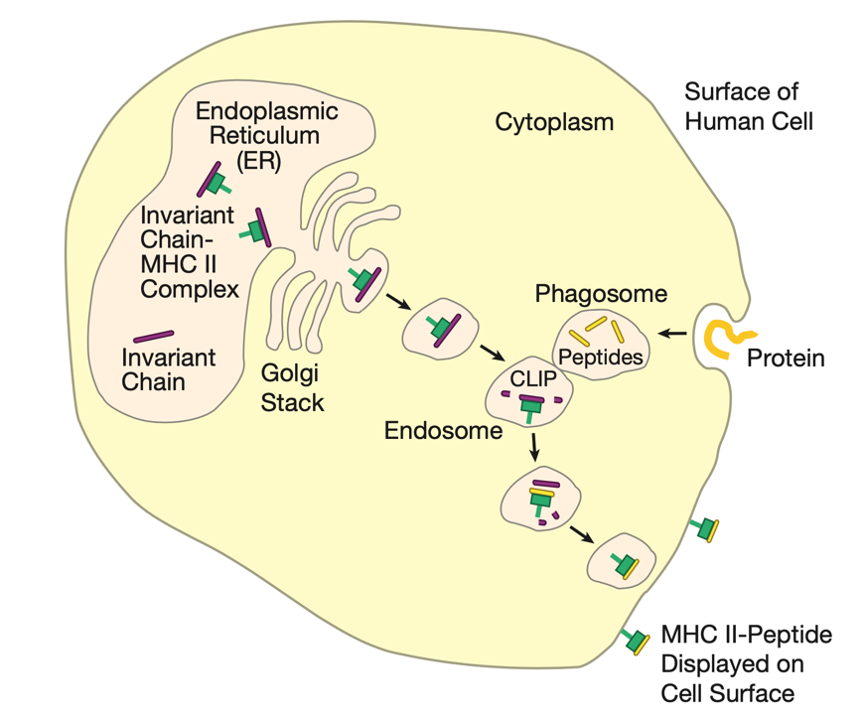
MHC II类分子介导的抗原提呈过程
组成MHC II类分子的α链与β链在胞质核糖体合成后,经信号肽引导进入内质网腔。在内质网中,新生的MHC II类分子异源二聚体与恒定链(Invariant chain, Ii)发生特异性结合。作为专职分子伴侣,恒定链通过以下机制实现抗原呈递的精准调控:
- 内源性肽段屏蔽功能:恒定链的CLIP片段(Class II-associated Invariant chain Peptide)占据MHC II类分子的抗原结合沟槽,形成物理屏障,阻止内质网腔内源性肽段(如蛋白酶体加工产生的短肽)非特异性结合。这种隔离机制避免了MHC I类与II类分子竞争性装载内源性抗原,确保两类分子分别执行内源性/外源性抗原监测的专职功能。
- 内体靶向运输功能:恒定链羧基端含有的内体分选信号(如LLN motif),引导MHC II类分子-恒定链复合体经高尔基体定向转运至内体-溶酶体区室,避开经典的分泌途径。
在内体-溶酶体系统的酸性环境中,恒定链经历逐步降解:首先由组织蛋白酶S(Cathepsin S)切除大部分序列,仅保留CLIP片段持续占据抗原结合沟。此时,外源性抗原加工进程同步启动:
- 抗原摄取与加工:细胞通过吞噬作用或受体介导的内吞作用摄取外源性蛋白,形成吞噬体。吞噬体与内体融合后,酸性水解酶(如组织蛋白酶B、L)将完整蛋白降解为13-25个氨基酸的抗原肽段。
- CLIP置换与肽段装载:HLA-DM分子(人源)/H2-M分子(鼠源)作为专职编辑酶,通过构象选择机制催化CLIP释放,同时稳定开放的MHC II类分子构象,促进高亲和力抗原肽段置换结合。此过程受肽段结合基序(如HLA-DR的锚定氨基酸偏好性)的严格筛选。
最终,成熟的MHC II类分子-抗原肽复合物经转运小泡递呈至细胞膜表面,形成TCR识别及CD4共受体结合的信号平台。
MHC I类与II类分子抗原加工系统的空间隔离机制
进化过程中,机体通过以下机制实现抗原呈递的分工协作:
- 定位隔离:MHC I类分子在内质网直接装载蛋白酶体产物,而MHC II类分子需经内体-溶酶体系统加工外源性抗原,形成物理隔绝。
- 分子伴侣调控:恒定链与TAP(抗原肽转运体)分别作为MHC II类与I类分子的专属伴侣,确保抗原装载的特异性。
- 免疫监视分工:MHC I类分子构成全身性监测网络,而MHC II类分子通过专职抗原呈递细胞的抽样检测,实现对外源性威胁的精准预警。这种分工策略在保障免疫广度的同时,避免了无效的免疫激活。
小结
在近期的系列解析中,我们系统阐述了MHC I类分子与MHC II类分子的分子特征及免疫学功能。尽管两类分子在亚细胞定位、抗原来源偏好及细胞分布模式上呈现显著差异,但其核心生物学功能均聚焦于抗原呈递——通过特异性识别并装载抗原肽段,形成T细胞受体(TCR)识别的分子界面。MHC I类分子凭借其广泛表达特性构建全身性免疫监视网络,实时检测细胞内蛋白合成异常;MHC II类分子则通过专职抗原呈递细胞的抽样检测机制,精准解析细胞外环境中的病原体相关分子模式。两类分子协同作用,共同构成适应性免疫应答的启动枢纽,其肽结合沟槽与抗原肽的共价结合及膜表面展示,奠定了T细胞介导免疫识别的分子基础。这种分工明确又相互补充的抗原呈递体系,是机体实现精准免疫防御与免疫稳态调控的关键机制。
相关商品推荐
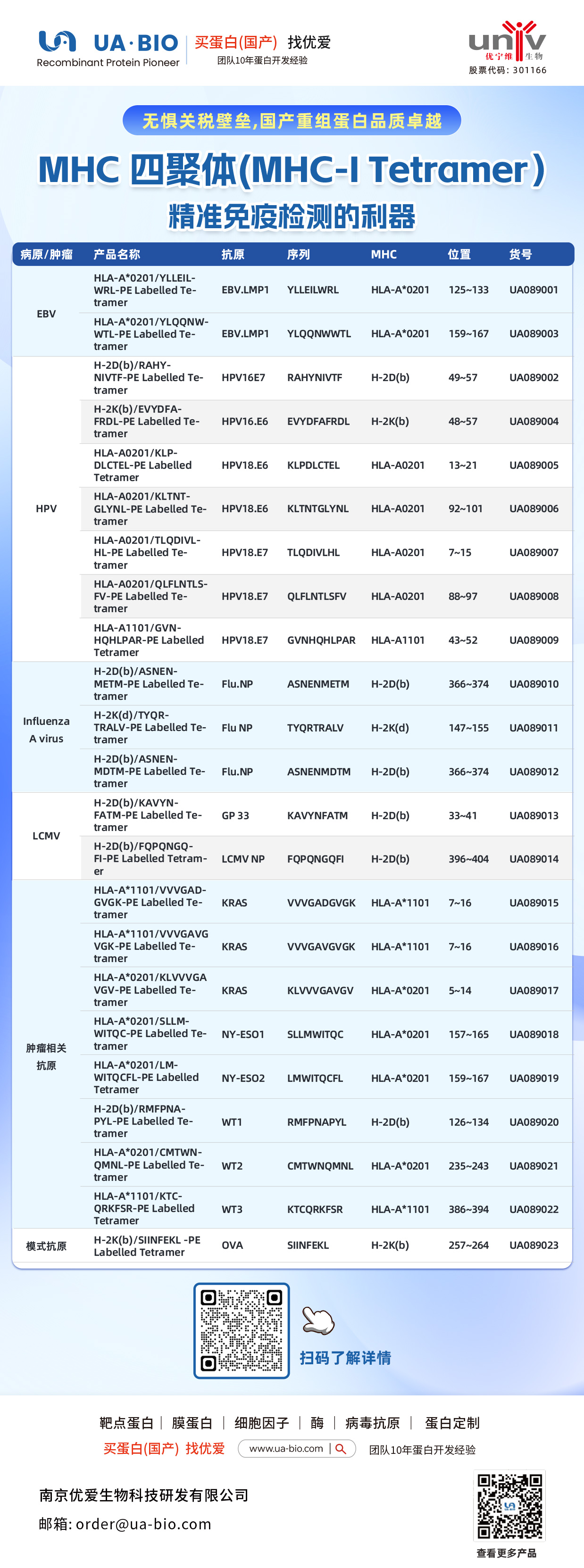
| 病原/肿瘤 | 产品名称 | 抗原 | 序列 | MHC | 位置 | 货号 |
| EBV | HLA-A*0201/YLELLVWRL-PE Labelled Tetramer | EBV.LMP1 | YLELLVWRL | HLA-A*0201 | 125-133 | UA089001 |
| EBV | HLA-A*0201/YLQQNWTL-PE Labelled Tetramer | EBV.LMP1 | YLQQNWTL | HLA-A*0201 | 159-167 | UA089003 |
| EBV | H-2Db(b)/RAHY-NIVTF-PE Labelled Tetramer | HPV16.E7 | RAHYNIVTF | H-2Db | 49-57 | UA089002 |
| HPV | H-2K(b)/EVYDFA-FRQL-PE Labelled Tetramer | HPV16.E6 | EVYDFARDL | H-2Kb | 48-57 | UA089004 |
| HPV | HLA-A*0201/KLP-DLCTL-PE Labelled Tetramer | HPV18.E6 | KLPDCTL | HLA-A*0201 | 13-21 | UA089005 |
| HPV | HLA-A*0201/KLTNT-GLYQL-PE Labelled Tetramer | HPV18.E6 | KLTNTGLYNL | HLA-A*0201 | 92-101 | UA089006 |
| HPV | HLA-A*0201/TLODIVIHL-PE Labelled Tetramer | HPV18.E7 | TLODIVIHL | HLA-A*0201 | 7~15 | UA089007 |
| HPV | HLA-A*0201/QFLNTL-FV-PE Labelled Tetramer | HPV18.E7 | QFLNTLFSV | HLA-A*0201 | 88-97 | UA089008 |
| HPV | HLA-A*1101/GVNHQLPAR-PE Labelled Tetramer | HPV18.E7 | GVNHQLPAR | HLA-A*1101 | 43-52 | UA089009 |
| Influenza A Virus | H-2D(b)/ASNENMETM-PE Labelled Tetramer | Flu.NP | ASNENMETM | H-2Db | 366-374 | UA089010 |
| Influenza A Virus | H-2K(d)/TYQR-TRALY-PE Labelled Tetramer | Flu.NP | TYQRTRALY | H-2Kd | 147-155 | UA089011 |
| Influenza A Virus | H-2D(b)/ASNEN-MDTM-PE Labelled Tetramer | Flu.NP | ASNENMDTM | H-2Db | 366-374 | UA089012 |
| LCMV | H-2D(b)/KAVYNFATM-PE Labelled Tetramer | GP 33 | KAVYNFATM | H-2Db | 33-41 | UA089013 |
| LCMV | H-2D(b)/FQPGQGFVK-PE Labelled Tetramer | LCMV NP | FQPGQGFVK | H-2Db | 396-404 | UA089014 |
| Tumor-related | HLA-A*1101/VVGADGVK-PE Labelled Tetramer | KRAS | VVGADGVK | HLA-A*1101 | 7~16 | UA089015 |
| Tumor-related | HLA-A*1101/VVGAGVGK-PE Labelled Tetramer | KRAS | VVGAGVGK | HLA-A*1101 | 7~16 | UA089016 |
| Tumor-related | HLA-A*0201/KLVVGAGV-PE Labelled Tetramer | KRAS | KLVVGAGV | HLA-A*0201 | 5~14 | UA089017 |
| Tumor-related | HLA-A*0201/SLLMWITQC-PE Labelled Tetramer | NY-ESO1 | SLLMWITQC | HLA-A*0201 | 157-165 | UA089018 |
| Melanoma | HLA-A*0201/LMWITQCFL-PE Labelled Tetramer | NY-ESO2 | LMWITQCFL | HLA-A*0201 | 159-167 | UA089019 |
| Melanoma | H-2Db(b)/MMFPNA-P1-PE Labelled Tetramer | WT1 | RMFPNAPL | H-2Db | 126-134 | UA089020 |
| Melanoma | HLA-A*0201/CMTWV-PE Labelled Tetramer | WT2 | CMTWVNMDM | HLA-A*0201 | 235-243 | UA089021 |
| Melanoma | HLA-A*1101/KTCQRKSF-PE Labelled Tetramer | WT3 | KTCQRKSF | HLA-A*1101 | 386-394 | UA089022 |
| Ovarian Cancer | H-2K(b)/SINFEKL-PE Labelled Tetramer | OVA | SINFEKL | H-2Kb | 257-264 | UA089023 |
声明:本篇文章在创作中部分采用了人工智能辅助。如有任何内容涉及版权或知识产权问题,敬请告知,我们承诺将在第一时间核实并撤下。
买重组蛋白,找南京优爱
优爱蛋白专注于提供药物研发、细胞治疗、基因治疗、基础科研所需各种蛋白类试剂原材料和服务,包括药物靶点蛋白、免疫检查点蛋白、细胞因子、工具酶、 蛋白定制表达、全长跨膜蛋白开发等。优爱致力于为客户提供优质的产品和专业服务,打造具有国际竞争力的高新技术企业。











