

- 最新进展
- 产品信息
最新进展
一、研究背景与科学问题
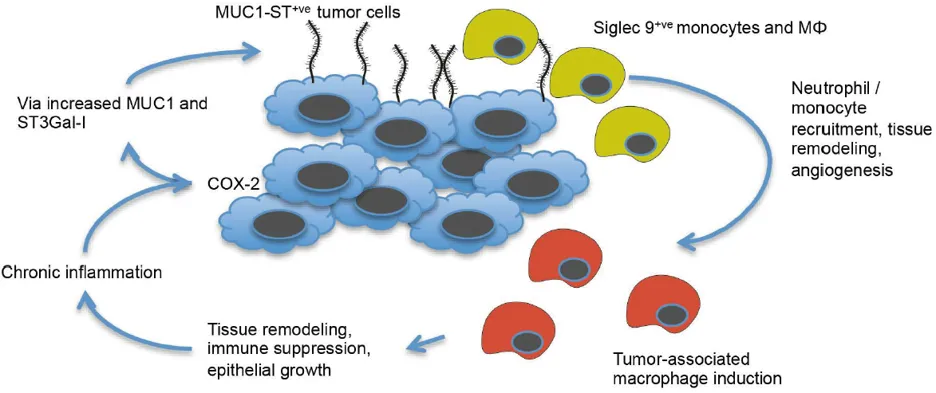
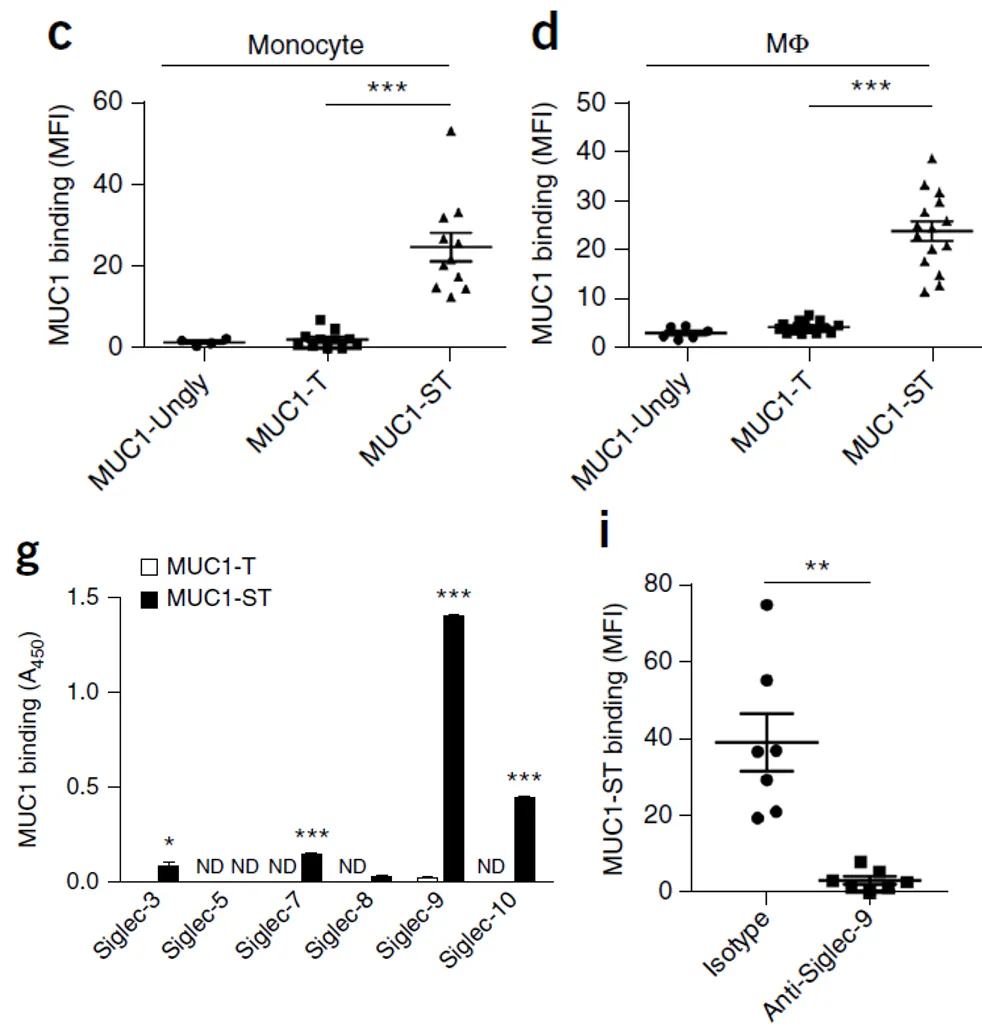
二、MUC1 异常唾液酸化与 Siglec-9 的特异性结合
MUC1-ungly:仅含蛋白骨架,无糖基化修饰;
MUC1-T:有基础糖基化,无唾液酸化;
MUC1-ST:同时具备糖基化与唾液酸化(异常修饰)。
三、Siglec-9 介导的单核细胞功能改变
促炎因子 IL-6 分泌增加,参与炎症微环境形成;
巨噬细胞集落刺激因子(MCSF)释放增多,促进单核细胞向巨噬细胞分化;
血管生成相关因子 PAI-1 上调,为肿瘤侵袭转移提供条件。
四、Siglec-9 依赖的巨噬细胞异常分化
表面共刺激分子 CD86 及 IL-12 表达显著下调;
对 CD8⁺T 细胞的增殖及激活分子 CD69 表达有显著抑制作用;
对树突状细胞(DC)的分化成熟也具有抑制作用。
五、巨噬细胞的促肿瘤表型诱导
分泌 M-CSF、PAI-1 及表皮生长因子(EGF)增加,参与巨噬细胞自我更新、肿瘤侵袭及细胞增殖调控;
高表达 M2 型巨噬细胞标志物 CD206、CD163;
免疫检查点分子 PD-L1 及 IDO 表达上调,强化免疫抑制。
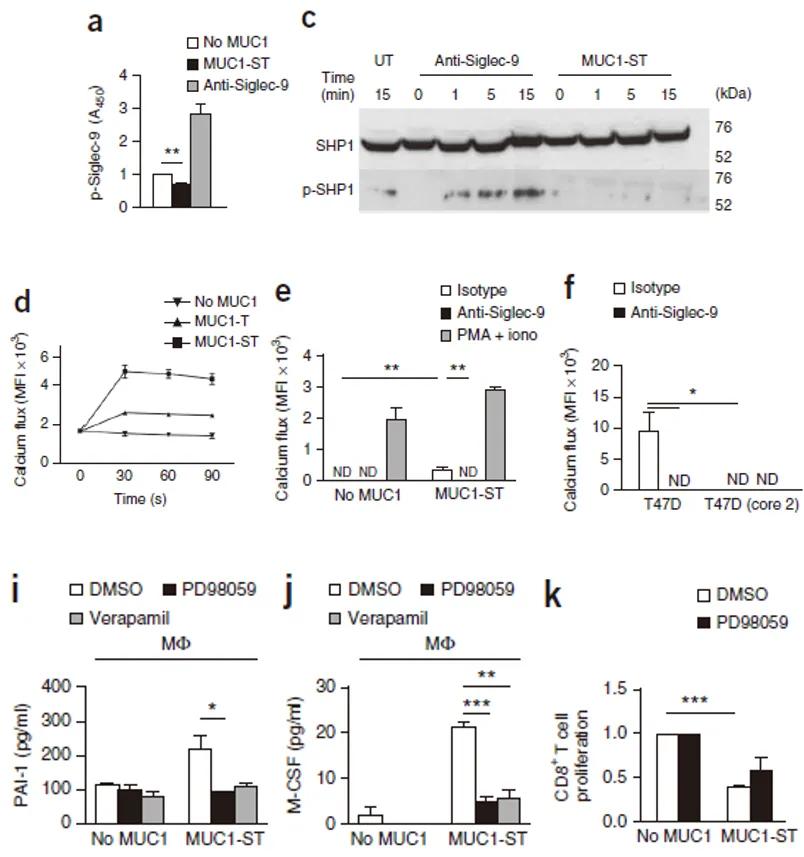
六、信号通路机制
显著增强巨噬细胞内钙离子转运,且该效应依赖 Siglec-9;
钙离子通道抑制剂维拉帕米可抑制促肿瘤因子分泌;
激活 MEK-ERK 通路,MEK 抑制剂可减少促肿瘤因子分泌,并部分恢复 CD8⁺T 细胞增殖能力。
七、研究意义
声明:本篇文章在创作中部分采用了人工智能辅助。如有任何内容涉及版权或知识产权问题,敬请告知,我们承诺将在第一时间核实并撤下。
买重组蛋白,找南京优爱
优爱蛋白专注于提供药物研发、细胞治疗、基因治疗、基础科研所需各种蛋白类试剂原材料和服务,包括药物靶点蛋白、免疫检查点蛋白、细胞因子、工具酶、 蛋白定制表达、全长跨膜蛋白开发等。优爱致力于为客户提供优质的产品和专业服务,打造具有国际竞争力的高新技术企业。
靶点蛋白 | 膜蛋白 | 细胞因子 | 酶 | 病毒抗原 | 蛋白定制
南京优爱生物科技研发有限公司 邮箱:order@ua-bio.com 热线:0571-87565022

产品信息

友情链接:









©2021-2026 南京优爱生物科技研发有限公司版权所有
网站备案号 浙ICP备2022019033号-2 浙公网安备33010202004867号
电话:0571-87565022
邮箱:order@ua-bio.com
本网站销售的所有产品均不得用于人类或动物之临床诊断或治疗,仅可用于工业或者科研等非医疗目的。
